Un regalo di Natale per L.U.C.A.

Caro L.U.C.A.,
Natale é già passato da quasi due settimane, ma trovo solo ora il tempo di mandarti una foto del regalo che ho deciso di farti per Natale! Come ben sai, tu sei l’ultimo antenato comune tra tutte le specie viventi, il nostro Last Universal Common Ancestor, e per questo ho deciso di regalarti un pesce! Mi prenderò cura di lui come se fosse un mio parente (anche se effettivamente lo é, perché risalendo i nostri alberi genealogici fino a 3,6/4 miliardi di anni fa dovremmo trovare entrambi te) e posterò regolarmente delle foto e dei dati sulla sua crescita.
Facendo delle ricerche sul tuo conto ho scoperto che in fondo, ma molto in fondo, ho dei legami, per quanto lontani, con tutti gli esseri viventi. Mi prenderò quindi cura non solo del pesce, a cui ho dato il nome Sushi, ma anche dell’ambiente in cui lui vivrà (sempre che vivrà, visto che, come puoi vedere dalla foto, il mio gatto lo sta già puntando) e mi preoccuperò di adattare l’acquario alle sue esigenze.
Ho infatti in programma di portare presto degli amici pesci a far compagnia a Sushi, magari prendendogli anche una nuova casa.
Nel frattempo terrò a bada il nemico!
Le sfere della Terra

Alcuni dei materiali che costituiscono la Terra si trovano allo stato solido, altri allo stato liquido e altri ancora allo stato aeriforme.
Basandoci sulla densità dei materiali possiamo distinguere tre sfere differenti
- idrosfera: componente liquida.
- atmosfera: componente gassosa.
- litosfera: componente solida superficiale.
La componente biologica dell’ambiente in cui viviamo è invece la biosfera.
Ecco le definizioni del enciclopedia Treccani delle varie sfere:
IDROSFERA L’involucro acqueo (mari, laghi, fiumi ecc.) che, con talune soluzioni di continuità rappresentate dalle terre emerse, copre la superficie della Terra per circa 361 milioni di km2 su un totale di 510 milioni. Per vari autori il termine comprende l’elemento acqueo presente sulla Terra anche sotto forma solida (ghiaccio) e aeriforme (vapore acqueo atmosferico).
ATMOSFERA Involucro gassoso che circonda o sovrasta un corpo solido o liquido; in particolare quello che circonda la Terra (a. terrestre) e altri pianeti (a. planetarie). Talora si parla di a. anche per il Sole e per le stelle, che sono corpi completamente gassosi: in tal caso s’intendono per a. gli strati più esterni e meno densi. La massa dell’a. terrestre è un milionesimo di quella della Terra; il 99% si trova al di sotto di 50 km di quota e il 90% al di sotto di 17 km.
LITOSFERA In geologia, lo strato reologico (soggetto a deformazioni) più esterno della Terra, caratterizzato da un comportamento rigido ed elastico. Si distingue sia una l. oceanica, costituita dall’insieme della crosta oceanica e dal mantello terrestre superiore, sia una l. continentale, formata a sua volta dalla crosta continentale e dal mantello terrestre superiore. Lo spessore della l. è variabile: sotto gli oceani esso si aggira mediamente sui 75 km, mentre sotto le aree continentali arriva fino a 110-130 km. Al di sotto delle dorsali oceaniche la l. presenta i minimi spessori, valutati intorno ai 5-10 km. La l. poggia su un substrato denominato astenosfera, che rappresenta, grazie alle temperature più elevate, una regione meno rigida della l. e che si deforma a causa del suo comportamento plastico. Gli sviluppi nel campo della geofisica hanno inoltre messo in evidenza che la l. non costituisce un involucro continuo sulla superficie terrestre, ma è rotta in numerosi punti lungo zone di frattura molto estese, sedi di intensa attività sismica. Queste zone delimitano un mosaico di zolle litosferiche che sono in moto relativo le une rispetto alle altre e che slittano sul substrato plastico della astenosfera
BIOSFERA Nome comprensivo per indicare quella parte della Terra nella quale si riscontrano le condizioni indispensabili alla vita animale e vegetale. Comprende la parte bassa dell’atmosfera, tutta l’idrosfera e la parte superficiale della litosfera, fino a 2 km di profondità. Insieme alle forme di vita che ospita, costituisce un sistema complesso, in equilibrio dinamico con le altre componenti della Terra.
La teoria della deriva dei continenti
Nel 1912 Alfred Wegener ideò la teoria della deriva dei continenti.
Ecco un video di RAIscienze che spiega questa teoria
Composizione interna del Pianeta Terra

Attraverso lo studio dei parametri che regolano la propagazione delle onde sismiche è possibile ricavare informazioni riguardanti lo stato fisico dei materiali costitutivi delle zone più interne della Terra e la loro relativa composizione.
Queste analisi hanno permesso di sviluppare un modello che descrive la struttura interna della Terra. La prima importante informazione sulla sua struttura riguarda la densità. La terra non ha una densità uniforme. Se le onde sismiche si propagassero lungo un gradiente di densità (densità che varia in modo costante) le loro curve non dovrebbero presentare grosse deviazioni. Invece è stato dimostrato (grazie all’analisi dei sismogrammi) che tali onde subiscono delle brusche deviazioni.
Cosa può giustificare tali deviazioni? Delle grosse discontinuità (e differenze di densità) dei vari strati che un’onda attraversa lungo il suo corso. (fonte qui)
Se si considera la strutturazione della Terra secondo il criterio chimico-mineralogico, la Terra è così composta:
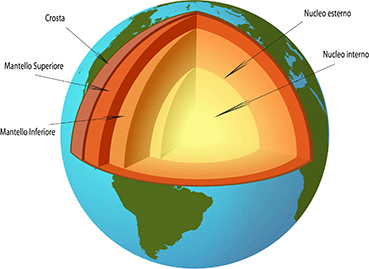
Basandosi invece sulla suddivisione degli involucri della Terra basata sullo stato fisico dei materiali, la Terra è fatta così:
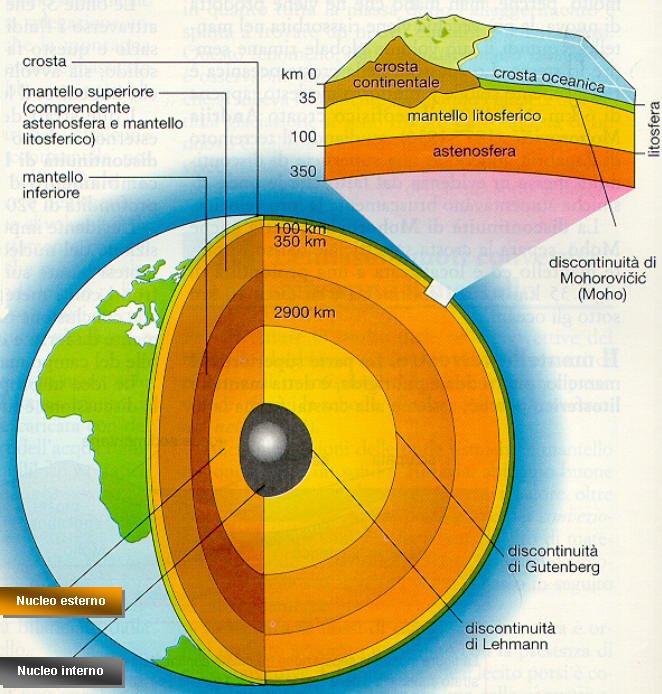
Le origini del Pianeta Terra

4,6 miliardi di anni fa l’agglomerazione di un insieme di polveri e gas dette origine al protosole e ai pianeti della Via Lattea, la nostra galassia. Tra questi pianeti vi era la Terra, le cui rocce più antiche risalgono a 3,8-4,2 miliardi di anni fa.
Il nostro pianeta si è formato a causa dell’agglomerazione di vari corpi celesti, i planetesimi, che urtarono la superficie. Mentre la Terra aumentava di volume, aumentava anche la sua temperatura.
Il surriscaldamento interno fu dovuto a tre principali motivi.
- accrescimento per impatti: con l’impatto dei planetesimi sulla superficie terrestre, la loro energia cinetica si trasformava in energia termica, in parta dispersa nello spazio e in parte trattenuta;
- compressione gravitativa: con l’aumento del volume del pianeta, e quindi il conseguente aumento del suo peso, aumentava anche la pressione sulle sue parti interne. Ciò comportava un notevole incremento di calore;
- disintegrazione di elementi radioattivi: le particelle atomiche rilasciate dagli elementi radioattivi, presenti in quantità nettamente maggiori rispetto ad adesso, vengono assorbite dall’ambiante circostante e la loro energia cinetica si trasforma in energia termica.
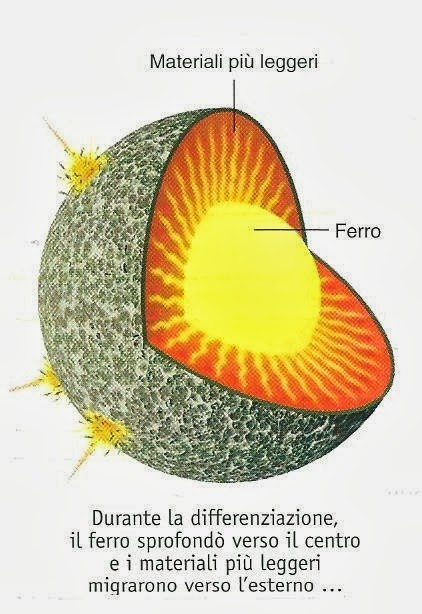
Per questi tre fenomeni la temperatura interna del pianeta raggiunse circa 1000 °C. A causa di questo continuo aumento di calore, dopo 300-1000 milioni di anni dall’origine della Terra si raggiunse la temperatura di fusione del ferro, dando origine a quel fenomeno denominato “catastrofe del ferro” (il ferro fuso, più denso, contenuto negli strati esterni della Terra, sprofondò verso il centro del pianeta e formò il nucleo).
Questo fenomeno determinò la riorganizzazione interna della Terra, che, da pianeta omogeneo, divenne un pianeta stratificato.
RECENSIONE LIBRO: Joyland di Stephen King

Quando mi é stato chiesto dal professore di scienze di consigliargli una piacevole lettura, ho subito avuto le idee chiare su cosa non avrei scelto: un libro di scienze. Non mi é stato invece chiaro fin da subito cosa consigliargli; lui aveva suggerito un libro di fantascienza, ma l’idea giusta mi é arrivata quando mia zia mi ha regalato un libro in occasione del mio compleanno. Il libro, Joyland appunto, non mi ha ispirato fin da subito. Solo quando ho iniziato a leggerlo, ho capito che era quello il romanzo giusto, un thriller.
Joyland, scritto da Stephen King e pubblicato il 4 giugno 2013 dalla Hard Case Crime, non é uno di quei thriller che ti tiene con il fiato sospeso fin dall’inizio, ma che ti fa appassionare un po’ alla volta alle vicende di Devin Jones, prima di entrare nel clou della vicenda.
Devin Jones é un normale studente universitario, che per guadagnare un po’ di soldi lavora nella mensa dell’istituto. Quando nel 1973 decide di lavorare a Joyland, un parco divertimenti della Carolina del Nord, per la stagione estiva, la ragazza che ama lo tradisce, spezzandogli il cuore e conducendolo anche a valutare più volte l’ipotesi del suicidio.
A salvarlo dal baratro sono i suoi amici conosciuti a Joyland, Tom ed Erin, che rappresenteranno molto per il protagonista, e anche dei dipendenti a tempo pieno del parco come Lane Hardy e Madame Fortuna, grazie ai quali viene a conoscenza del mistero che grava sul l’attrazione più spaventosa del parco: il Castello del Brivido. Pare infatti che in esso si nasconda il fantasma di una ragazza assassinata nel buio dell’attrazione dal suo fidanzato, che non ha lasciato nessuna traccia oltre alla fotografie, scattate dalle sirene di Hollywood, dipendenti del parco, che lo avevano ritratto in compagnia della fanciulla e che mostravano alcuni suoi segni particolari: capelli biondi e un tatuaggio a forma di aquila sul polso. Personaggi chiave per la storia sono anche Mike, un ragazzino che soffre di distrofia di Duchenne, destinato a morire, e Annie Ross, sua madre, di cui Devin finirà per innamorarsi.
Un libro carino, appassionante e a tratti provocante. Peccato per un finale da cui ci si aspettava certamente di più, una delle poche pecche di quest’opera King.
VALUTAZIONE: 4/5
RECENSIONE BLOG: Keplero
“Keplero è il blog di Amedeo Balbi. Divulgazione scientifica (astrofisica e cosmologia, ma non solo) con un occhio alla cultura pop, e (sporadiche) divagazioni personali.”
Un blog veramente ben fatto, curato e costantemente aggiornato.
Questo é per me Keplero. Amedeo Balbi ha fatto, e sta facendo, veramente un buon lavoro. Sul suo sito ci sono tanti curiosi articoli, i cui argomenti spaziano dalla chimica alla biologia, dalla fisica alla matematica, dalla cultura generale alla musica. Durante l’estate la sua produzione si é arrestata, piccolo particolare che si può perdonare visto il ritmo con cui a ripreso: dall’inizio di settembre ad oggi ha scritto 6 articoli, uno ogni due giorni.
I suoi post non sono molto lunghi, anzi, a volte si limita a scrivere poche righe, una pecca nel suo lavoro. Spesso allega dei video ai suoi articoli, grazie ai quali si risparmia una buona dose di lavoro, ma soprattutto risparmia parecchio impegno ai lettori meno volenterosi. Gli argomenti sono forse troppo vari: un particolare tema non viene quasi mai approfondito o ripreso da altri post, perdendo appeal tra coloro che preferiscono i blog che seguono sempre la stessa materia.
Complessivamente il blog é buono e utilizza un linguaggio di facile comprensione per chiunque, anche se a volte gli argomenti vengono trattati con troppa leggerezza.
VALUTAZIONE: 3.5/5
Un nuovo inizio
E così anche queste vacanze estive sono terminate e sta per iniziare un nuovo anno scolastico. Da oggi si ricomincia a scrivere sul blog e a studiare.
Auguro a tutti un anno fantastico e pieno di soddisfazioni!!! Mi raccomando, non fatevi bocciare.
Population Genetics: When Darwin Met Mendel

Ogni volta che visito il canale youtube crashcourse trovo sempre qualcosa di interessante!
Oggi mi sono imbattuto in un video in cui Gregor Mendel incontra Charles Darwin, con qualche accenno all’appena trattato equilibrio di Hardy-Weinberg.
L’equilibrio di Hardy-Weinberg
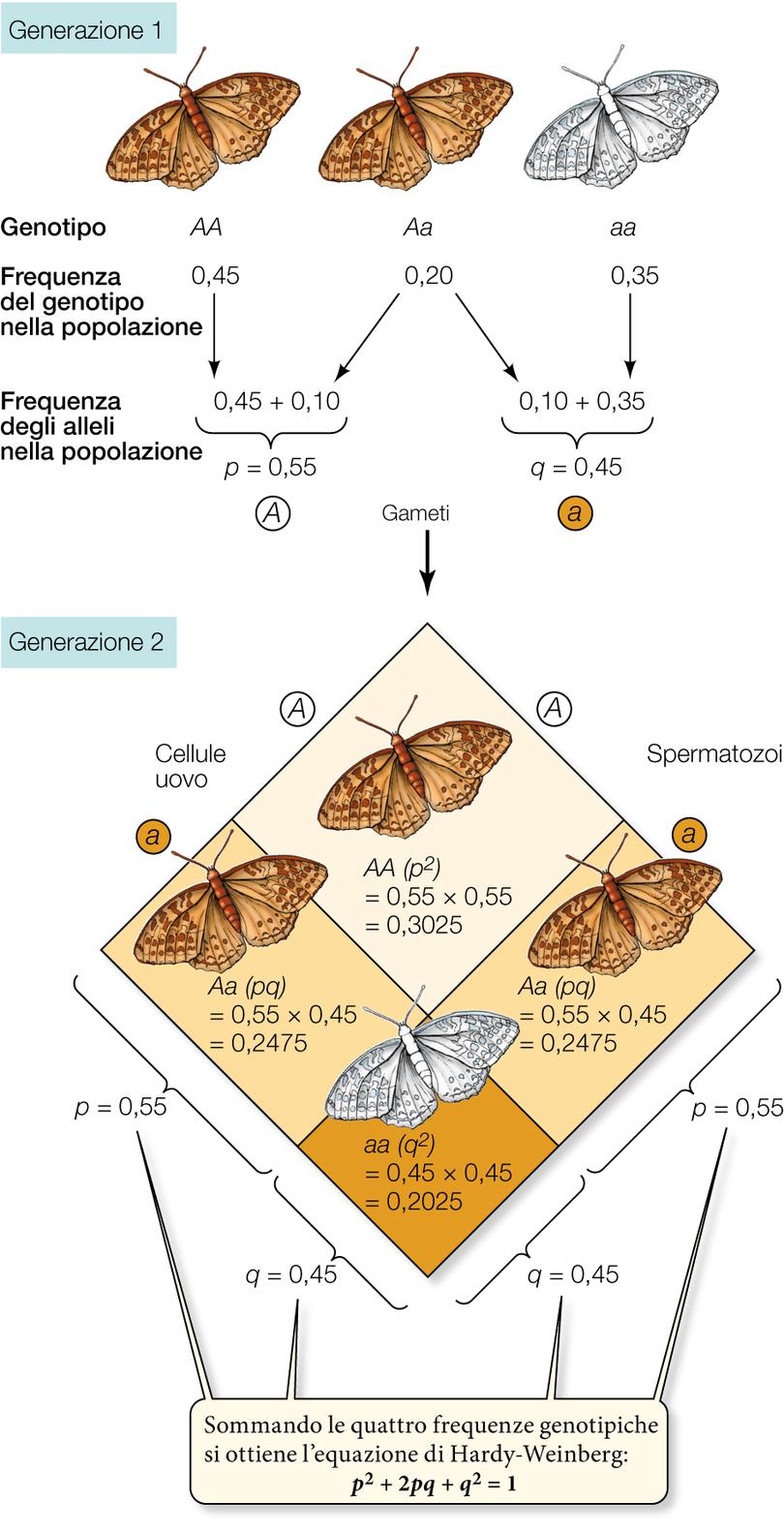
Nel 1908, il matematico britannico Godfrey Hardy e il medico tedesco Wilhelm Weinberg dedussero le condizioni necessarie perché la struttura genetica di una popolazione si mantenga invariata nel tempo.
Il concetto di equilibrio di Hardy-Weinberg è la chiave di volta della genetica di popolazione. La relativa equazione descrive una situazione modello in cui le frequenze alleliche rimangono costanti da una generazione all’altra e le frequenze genotipiche sono ricavabili da quelle alleliche. La legge dell’equilibrio di Hardy-Weinberg si applica agli organismi che si riproducono sessualmente. Le condizioni che devono essere soddisfatte affinché una popolazione si trovi all’equilibrio di Hardy-Weinberg sono le seguenti:
- Gli accoppiamenti devono essere casuali.
- Gli individui non devono preferire partner con particolari genotipi.
- La popolazione deve essere di grandi dimensioni.
- Più grande è la popolazione, minore è l’effetto delle eventuali fluttuazioni casuali delle frequenze alleliche.
- Non deve esserci flusso genico.
- In altre parole, non devono verificarsi fenomeni di immigrazione o di emigrazione.
- Non devono avvenire mutazioni.
- Gli alleli non si trasformano uno nell’altro né possono comparirne di nuovi.
- La selezione naturale non deve influenzare la sopravvivenza di particolari genotipi.
- Gli individui con genotipi diversi hanno la stessa possibilità di sopravvivere.
Se queste condizioni sono idealmente soddisfatte, ne seguono due importanti conseguenze. Primo, dopo una generazione di accoppiamenti casuali, se p è la frequenza allelica di A e q è la frequenza allelica di a, le frequenze genotipiche manterranno i seguenti rapporti:
| genotipo | AA |
Aa |
aa |
| frequenza | p2 |
2pq |
q2 |
Considera la generazione 1 in cui la frequenza dell’allele A (p) è 0,55. Poiché abbiamo ipotizzato che gli individui scelgano i propri partner casualmente, senza considerare il loro genotipo, i gameti portatori dell’allele A oppure dell’allele a si combinano casualmente, cioè secondo quanto previsto dalle rispettive frequenze p e q.
Nel nostro esempio, la probabilità che un particolare gamete porti un allele A anziché a è di 0,55. In altre parole, su 100 gameti presi a caso, 55 recheranno l’allele A. Dato che q = 1 − p, la probabilità che uno spermatozoo o una cellula uovo rechi l’allele a sarà 1 − 0,55 = 0,45.
La probabilità che alla fecondazione l’incontro avvenga tra due gameti portatori di A è data dal prodotto delle due probabilità relative ai singoli eventi:
p x p = p2 = (0,55)2 = 0,3025
Quindi, nella generazione successiva, il 30,25% della prole avrà genotipo AA. Allo stesso modo, la probabilità che si incontrino due gameti portatori di a sarà
q x q = q2 = (0,45)2 = 0,2025
e il 20,25% della generazione successiva avrà genotipo aa.
La seconda conseguenza è che le frequenze p e q degli alleli di un gene rimangono costanti di generazione in generazione, come ora è facile dimostrare. Infatti nella nuova generazione della nostra popolazione ad accoppiamenti casuali la frequenza dell’allele A è p2 + pq, e sostituendo q con 1 − p, l’espressione diventa:
p2 + p (1 − p) = p2 + p – p2 = p
Le frequenze alleliche di partenza restano immutate, e la popolazione si trova all’equilibrio, espresso dall’equazione di Hardy-Weinberg:
p2+ 2pq + q2= 1
Se le frequenze genotipiche nella generazione parentale dovessero cambiare (per esempio, per l’emigrazione di un gran numero di individui AA), anche le frequenze alleliche nella generazione successiva risulterebbero alterate. Tuttavia, partendo dalle nuove frequenze alleliche, basta una sola generazione prodotta in seguito ad accoppiamenti casuali per riportare le frequenze genotipiche all’equilibrio.
(Tratto liberamente da http://ebook.scuola.zanichelli.it/sadavabiologia/l-evoluzione-dopo-darwin-la-teoria-sintetica/document-85)
Ecco anche un trattato di 12 pagine che spiega dettagliatamente LA LEGGE DI HARDY-WEINBERG
H. J. Muller

Hermann Joseph Muller scoprì che alcune radiazioni possono avere l’effetto di mutageni. Mendel aveva dimostrato che le coppie di alleli segregano indipendentemente dalle altre coppie. In realtà gli alleli di due geni differenti possono segregare indipendentemente solo se i geni sono su due cromosomi diversi. I geni che tendono a rimanere insieme perché appartenenti allo stesso cromosoma appartengono allo stesso gruppo di associazione.
Quando furono incrociati individui con il corpo marrone chiaro e le ali lunghe con i mutanti con corpo nero e ali corte, nacque una generazione F1 con le caratteristiche normali.
Dall’incrocio tra gli individui F1, si ipotizzò che o i geni per il colore del corpo e la lunghezza delle ali si sarebbero potuti segregare indipendentemente generando un rapporto 9:3:3:1, indicando che i geni per i due caratteri si trovavano su coppie differenti di cromosomi omologhi, o che i geni potevano essere associati e sarebbero nati il 75 % marrone con ali lunghe e il 25 % nero con ali corte. I risultati si avvicinarono alla seconda ipotesi ma alcuni geni segregavano indipendentemente.
Morgan suppose che ci potesse essere uno scambio di alleli tra cromosomi omologhi, il crossing over.
“Il crossing over è l’importante meccanismo di ricombinazione del materiale genetico proveniente dai due genitori, che permette una maggiore varietà nei prodotti della riproduzione sessuata.“ (Definizione di crossing over, tratta da Wikipedia)
Thomas Hunt Morgan

Morgan fu il primo ad effettuare degli studi sui caratteri trasportati da geni portati da cromosomi legati al sesso.
Compì i suoi studi sulla Drosophila melanogaster, il moscerino della frutta che possiede solo quattro cromosomi. Quando un giorno nella colonia di moscerini dagli occhi rossi ne comparve uno con gli occhi bianchi (maschio mutante), lo fece incrociare con una femmina con occhi rossi e in tutta la generazione F1 erano presenti solo moscerini con occhi rossi, facendo supporre che il fenotipo fosse recessivo.

Morgan incrociò allora gli individui della generazione F1 tra di loro, ma non si manifestò il rapporto 3:1 tra dominante e recessivo che si attendeva. Per di più erano solo maschi gli individui con gli occhi bianchi. Morgan incrociò quindi il primo maschio occhi bianchi con una femmina della generazione F1 eterozigote.
Dai risultati ottenuti dedusse che il gene per il colore degli occhi é presente solo sul cromosoma X e che l’allele per il carattere occhi bianchi dovesse essere recessivo.
In seguito ai risultati ottenuti Morgan non solo confermò che sono i cromosomi a trasportare i geni, ma anche che alcuni geni possono essere trasportati solo da cromosomi legati al sesso (X e Y).
Walter Sutton e i cromosomi

Nel 1902 Walter Sutton stava studiando la produzione di gameti nei maschi della cavalletta, quando notò che i cromosomi risultavano appaiati sin dall’inizio della prima divisione meiotica e i due cromosomi di ogni coppia erano molto simili.
Sutton fu colpito dal parallelismo delle sue osservazioni con la legge di Mendel sulla segregazione*, ipotizzando che i geni si trovassero nei cromosomi e che i due alleli di ogni gene si trovassero su cromosomi omologhi. Supponendo inoltre che gli alleli fossero sempre indipendenti e fossero separati nella meiosi I, dedusse che, durante la fecondazione, potessero nascere nuove combinazioni di alleli.
Riguardo la terza legge di Mendel** (gli alleli di geni diversi si separano indipendentemente gli uni con gli altri) sostenne la sua valida, purché i geni non si trovassero sulla stessa coppia di cromosomi omologhi.
Il suo principale contributo alla biologia fu quindi l’aver messo in relazione l’ereditarietà con i cromosomi a livello cellulare.
*SEGREGAZIONE: ogni individuo possiede coppie di fattori per ogni unità ereditaria e i membri di una coppia si separano (segregano) nella formazione dei gameti.
**ASSORTIMENTO INDIPENDENTE: dall’incrocio di due eterozigoti della generazione F1, si ottiene una generazione F2 in cui i caratteri segregano in maniera del tutto indipendente, originando nuove combinazione che seguono proporzioni definite.
Correlazione genotipo-fenotipo
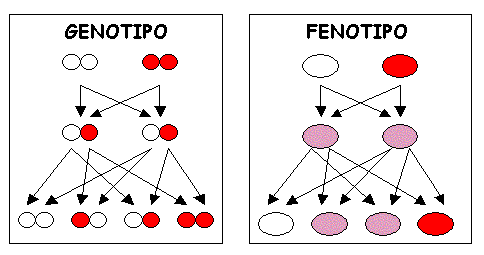
Ecco alcune definizioni che possono essere utili per ripassare gli effetti dell’interazione fra i geni sul fenotipo:
DOMINANZA INCOMPLETA: il fenotipo dell’eterozigote presenta caratteristiche intermedie tra quelle dei due omozigoti.
CODOMINANZA: organismi eterozigoti non presentano caratteristiche intermedie ma mostrano entrambi i fenotipi eterozigoti.
All’interno di una popolazione di organismi é possibile che ci siano più di due forme alleliche per uno stesso gene. Si tratta di ALLELI MULTIPLI derivanti da più mutazioni di uno stesso gene.
Le caratteristiche del fenotipo di un organismo sono per lo più derivanti dall’interazione tra due o più geni distinti; in questo caso possono apparire dei fenotipi completamente nuovi.
In altri casi possono non essere prodotti dei nuovi fenotipi, ma un gene maschera gli effetti di un altro (EPISTASI).
É detto EREDITA’ POLIGENICA quel fenomeno che coinvolge più di due geni nella determinazioni dei caratteri dell’individuo. In questo caso il carattere presenta una gradazione di lievi differenze detta variazione continua.
PLEIOTROPIA: quando un singolo gene ha molteplici effetti sul fenotipo di un organismo.
Hugo de Vries

Hugo De Vries studiò le mutazioni su una pianta di rapunzia a grandi fiori, ipotizzando che esse causassero la comparsa di nuovi caratteri del tutto assenti nelle generazioni precedenti.
Elaborò così la teoria delle mutazioni, secondo la quale ogni organismo vivente nel corso della sua storia subisce un certo numero di mutazioni che tramanda alla sua discendenza. Inizialmente questa teoria entrò in contrasto con quella darwiniana, non conciliando la lenta evoluzione ipotizzata da Charles Darwin con i cambiamenti repentini provocati dalle mutazioni osservate da de Vries, solo successivamente si riuscì a dimostrare la validità di entrambe le tesi.
De Vries è noto anche per aver riscoperto la teoria di Mendel.
Negli ultimi anni le teorie di De Vries sono state ampiamente rivisitate: ad oggi si ritiene errato parlare di mutazioni com da lui intese (macromutazioni), ma si deve piuttosto parlare di micromutazioni.
Le tre leggi di Mendel
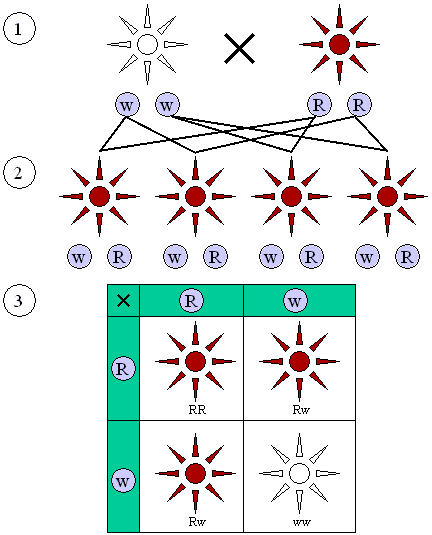
Frate Gregor non fu solo geniale, ma anche innovativo. Per giungere alle sue straordinarie scoperte, egli organizzò il suo lavoro in modo che potesse raggiungere risultati più precisi e dati più ricchi e facilmente comprensibili.
Egli strutturò la ricerca in questo modo:
-ipotesi di lavoro;
-pianificazione ben curata e scelta di studiare solo caratteristiche ereditarie nette;
-studio dei discendenti di più generazioni;
-conteggio del numero dei discendenti e analisi dei risultati.
Questo nuovo metodo lo portò all’elaborazione delle tre leggi che stanno alla base della genetica classica.
Mendel compì i suoi studi sulle piante di pisello, delle quali aveva identificato sette linee pure.
Dopo l’incrocio tra due linee pure nacque la prima generazione F1, che presentava solo uno dei due caratteri presente nei genitori (dominante); l’altro venne chiamato recessivo.
DOMINANZA: dall’incrocio di organismi che differiscono per una coppia di caratteri si ottengono solo individui che mostrano il carattere dominante.
Mendel notò che nella seconda generazione, nata dall’incrocio tra individui della prima generazione, i caratteri dominanti si presentavano in rapporto 3:1.
SEGREGAZIONE: ogni individuo possiede coppie di fattori per ogni unità ereditaria e i membri di una coppia si separano (segregano) nella formazione dei gameti.
Incrociando infine gli eterozigoti della generazione F1, che differivano tra di loro per due o più caratteri, osservò che i fenotipi della prole si presentavano in rapporto 9:3:3:1.
ASSORTIMENTO INDIPENDENTE: dall’incrocio di due eterozigoti della generazione F1, si ottiene una generazione F2 in cui i caratteri segregano in maniera del tutto indipendente, originando nuove combinazione che seguono proporzioni definite.
Quell’imbroglione di Mendel
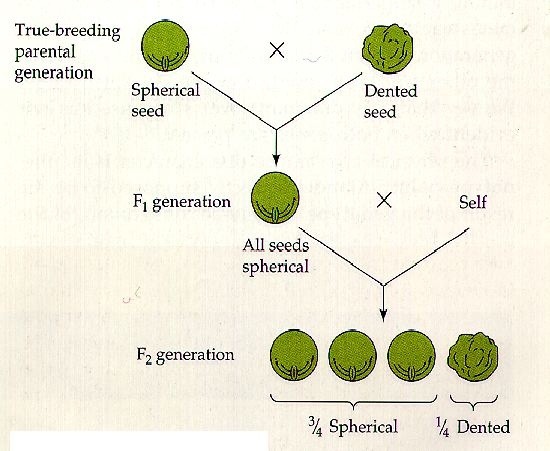
Non tutti sanno che il geniale frate Gregor Mendel, per divenire quello che noi ora riteniamo il padre della genetica, imbrogliò. Viste le conoscenze e gli strumenti a disposizione del naturalista in quella lontana epoca storica, si ritiene sia impossibile, o comunque alquanto improbabile, che con così pochi dati da prendere in considerazione, i risultati rispecchiassero alla perfezione la sua teoria.
Ma non concentriamoci su come arrivò alle sue leggi, bensì su come quest’ultime sono e che cosa comportano.
Ecco una dettagliata spiegazione di come si svolse il lavoro del frate agostiniano.
Le mutazioni geniche

Per mutazione genica si intende quella variazione, anche minima, che avviene nella struttura molecolare di un gene e che può riguardare un solo nucleotide, come anche più nucleotidi, della catena di DNA, come ad esempio la sostituzione di una purina o di una pirimidina con un’altra base azotata (per es.: come avviene nell’anemia falciforme si ha la sostituzione della timina con l’adenina). Una volta avvenuta questa mutazione nella struttura molecolare del materiale genico, essa si riproduce a ogni replicazione del DNA e cioè diventa ereditaria; soltanto un altro cambiamento potrà produrre una nuova mutazione che potrebbe anche consistere in un ritorno alla struttura originale.
Sono, quindi, le mutazioni che alterano un singolo gene e dunque le più “piccole” che si possono avere. In quanto tali non sono visibili attraverso analisi al microscopio (tranne alcuni casi estremi), ma possono essere riscontrate solo tramite analisi genetiche. Le mutazioni geniche portano alla formazione di nuove forme geniche, ovvero di nuovi alleli, detti appunto alleli mutanti. In quanto tali questi sono rari nella popolazione e si differenziano dagli alleli più diffusi detti invece tipi selvatici. Bisogna però far distinzione anche tra alleli mutanti e morfi. I morfi sono infatti due o più alleli di uno stesso gene con frequenza superiore all’1% (polimorfismo). Alla luce di questo ne deriva che il concetto di mutazione non è assoluto: un gene potrà subire una mutazione; se l’allele mutante però troverà le condizioni per diffondersi nella popolazione e superare la frequenza dell’1% non si parlerà più di mutazione ma di morfo.
Possono essere distinte in due categorie: mutazioni puntiformi e mutazioni per sequenze ripetute. Le prime sono causate da sostituzioni di basi o da inserzioni o delezioni di coppie di basi (mutazioni indel). La seconda categoria comprende le mutazioni causate sempre da inserzioni o delezioni ma di sequenze di basi ripetute.
Mutazioni puntiformi
Sostituzioni di basi: Determinano uno scambio di un nucleotide con un altro. Sono definite transizioni qualora vi è un scambio di una purina con altra purina (A ↔ G) o di una pirimidina con un’altra pirimidina (C ↔ T); oppure transversioni quando lo scambio è di una purina con un a pirimidina o viceversa (C/T ↔ A/G). In genere le transizioni sono più frequenti delle transversioni. Quando ci si riferisce a mutazioni di una sequenza che codifica per un determinato prodotto genico le sostituzioni potranno essere:
Mutazioni per sequenze ripetute
Analoghe alle mutazioni indel, interessano però più di un nucleotide adiacente; in particolare interessano gruppi nucleotidici che formano una sequenza la quale si ripete più volte di seguito. La mutazione, che si origina nel corso della replicazione del DNA, provoca una variazione nel numero di queste sequenze ripetute; il nuovo filamento di DNA potrà presentarne in eccesso o in difetto. Il fenomeno che causa la mutazione è detto slittamento della replicazione (replication slippage). Malattie genetiche associate a questo tipo di mutazione sono la Corea di Huntington e la sindrome dell’X fragile.
Effetti delle mutazioni geniche
Gli effetti possono essere notevolmente diversi a seconda del tipo di mutazione e della posizione in cui questa si verifica. Una mutazione può non portare a nessuna conseguenza e questo quando interessa DNA che non codifica (o meglio sembra non codificare) per nessun prodotto genico (il cosiddetto junk DNA o DNA spazzatura). Se la mutazione va invece ad alterare le sequenze codificanti, ovvero i geni, si ha una variazione nel tipo o nella quantità del corrispettivo prodotto genico, che può essere una proteina o RNA funzionale (rRNA, tRNA, snRNA ecc.). Parliamo in questo caso di mutazione biochimica; se la mutazione biochimica porta a una variazione visibile del fenotipo si parla di mutazione morfologica.
Inoltre distinguiamo, sempre in relazione agli effetti, in:
mutazione positiva: quella che porta un vantaggio evolutivo;
mutazione neutra: quella che non risulta in un depotenziamento della capacità riproduttiva dell’individuo;
mutazione subletale: quella che rende più difficoltosa la perpetuazione riproduttiva dell’individuo (il tipico esempio sono le malattie genetiche che debilitano in qualche modo l’individuo, rendendolo meno capace di riprodursi, senza però impedirglielo totalmente);
mutazione letale: quella che non permette all’individuo di raggiungere l’età riproduttiva o non gli permette di riprodursi.
L’efficacia della mutazione, sia positiva che negativa, dipende poi dal tipo di allele mutato così creato; questo potrà essere infatti dominante o recessivo. Nei diploidi (2n cromosomi) se è dominante avrà sempre effetto (sia in un eterozigote che in un omozigote dominante); se è recessivo, essendo aploinsufficiente, per avere effetto ha bisogno che anche l’altro elemento della coppia genica sia mutato (individuo omozigote recessivo). Negli aploidi , che sono emizigoti (n cromosomi), la mutazione avrà invece sempre effetto. (Fonte adattata e rielaborata: ftp://89-97-218-226.ip19.fastwebnet.it/web1/DNA/dna.htm)
Dal minuto 4.47 del video, un esempio di quotidiana mutazione genica:
La sintesi proteica
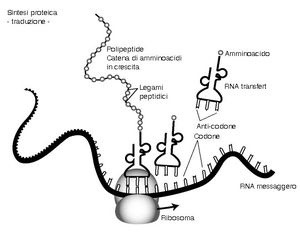
La traduzione è lo stadio della sintesi proteica in cui le istruzioni portate dall’m-RNA vengono tradotte nella sequenza corretta di amminoacidi per formare una proteina.
La traduzione ha luogo nel ribosoma (formato da r-RNA e proteine), composto da due subunità: quella piccola contiene un sito di legame per l’m-RNA; quella grande ha due siti di legame per due molecole di t-RNA e un sito che catalizza la formazione del legame peptidico tra due amminoacidi adiacenti.
Ogni molecola di t-RNA è specifica per un unico amminoacido ed è in grado di riconoscere sia l’amminoacido che deve trasportare, sia il codone complementare di m-RNA associato al ribosoma.
La traduzione ha inizio quando due codoni del filamento di m-RNA si legano alla subunità piccola di un ribosoma. Il primo codone è la tripletta di “inizio lettura” AUG, alla quale corrisponde l’amminoacido metionina; il secondo codifica il primo vero amminoacido della proteina. I due t-RNA, che hanno rispettivamente l’anticodone di inizio e l’anticodone complementare al secondo codone, si legano alla subunità grande e si forma un legame peptidico (cioè il legame tra amminoacidi che forma le proteine) tra i due amminoacidi trasportati.
Il t-RNA di inizio si stacca dal ribosoma mentre il dipeptide (i due amminoacidi uniti dal legame peptidico) rimane legato al secondo t-RNA. Il ribosoma si sposta sopra un altro codone dell’m-RNA e una nuova molecola di t-RNA con il proprio amminoacido si dispone nel sito di legame vuoto del ribosoma. Si crea un nuovo legame peptidico e il tripeptide si salda all’ultimo t-RNA. Il processo di allungamento della catena polipeptidica prosegue in questo modo finché tutte le triplette sono state tradotte e viene raggiunto il codone di “fine lettura”. La proteina completa si stacca dal ribosoma e specifici enzimi scindono il legame con la metionina.
Ecco inoltre un video esemplificativo dell’argomento trattato:
Il codice genetico

Come ben sappiamo ogni cromosoma é formato sia da proteine che da DNA. Ogni proteina contiene 20 amminoacidi differenti, ma il DNA e l’RNA, fonti del codice genetico delle proteine, contengono solo quattro basi azotate. Come é possibile?
Se ciascun nucleotide codificasse per un amminoacido, alle quattro basi corrisponderebbero solo quattro amminoacidi. Analogamente, se un amminoacido fosse codificato da due nucleotidi ci sarebbero al massimo 16 amminoacidi, comunque non sufficienti. Per ottenere un numero più che sufficiente di combinazioni, ogni amminoacido deve essere quindi codificato da tre nucleotidi, per un totale di 64 amminoacidi. Ogni combinazione é quindi costituita da una sequenza di tre nucleotidi chiamata codone.
Di queste 64 combinazioni, 3 rappresentano segnali di arresto (UUA, UAG e UGA) e le restanti 61 dei veri e propri amminoacidi, corrispondenti ai soli 20 presenti nella proteina. Ciò é possibile poiché i codoni ai quali corrisponde uno stesso amminoacido differiscono spesso solo per il terzo nucleotide ed é per questo che il codice viene definito degenerato.
